
מזרח אגן הים־התיכון מתאפיין בפסיפס גאולוגי, טופוגרפי ואקלימי היוצר מארג של בתי גידול המשתלבים זה בזה ומפגיש מינים (species) ממוצא טרופי, יובשני וממוזג. היות שגמלי־שלמה במוצאם האבולוציוני הם יצורים תֶּרְמוֹפִילִיִּים (חובבי חום), אין זה מפתיע שעושר המינים בפאונת גמלי־השלמה של ישראל – המונה כ־33 מינים – מתרכז באזורים החמים והיובשניים של הארץ (Simon et al., 2025).
מרבית המינים של גמלי־השלמה במרחב אגן הים־תיכון מייצגים פאונה שמוצאה טרופי וטרופי־יובשני, מאפריקה ומאסיה; כלומר, האבות הקדמונים שלהם התפתחו הרחק מכאן, ורוב המינים בסוגים (genera) או בשבטים (tribes) שאליהם הם משתייכים עדיין מצויים באזורי המוצא.
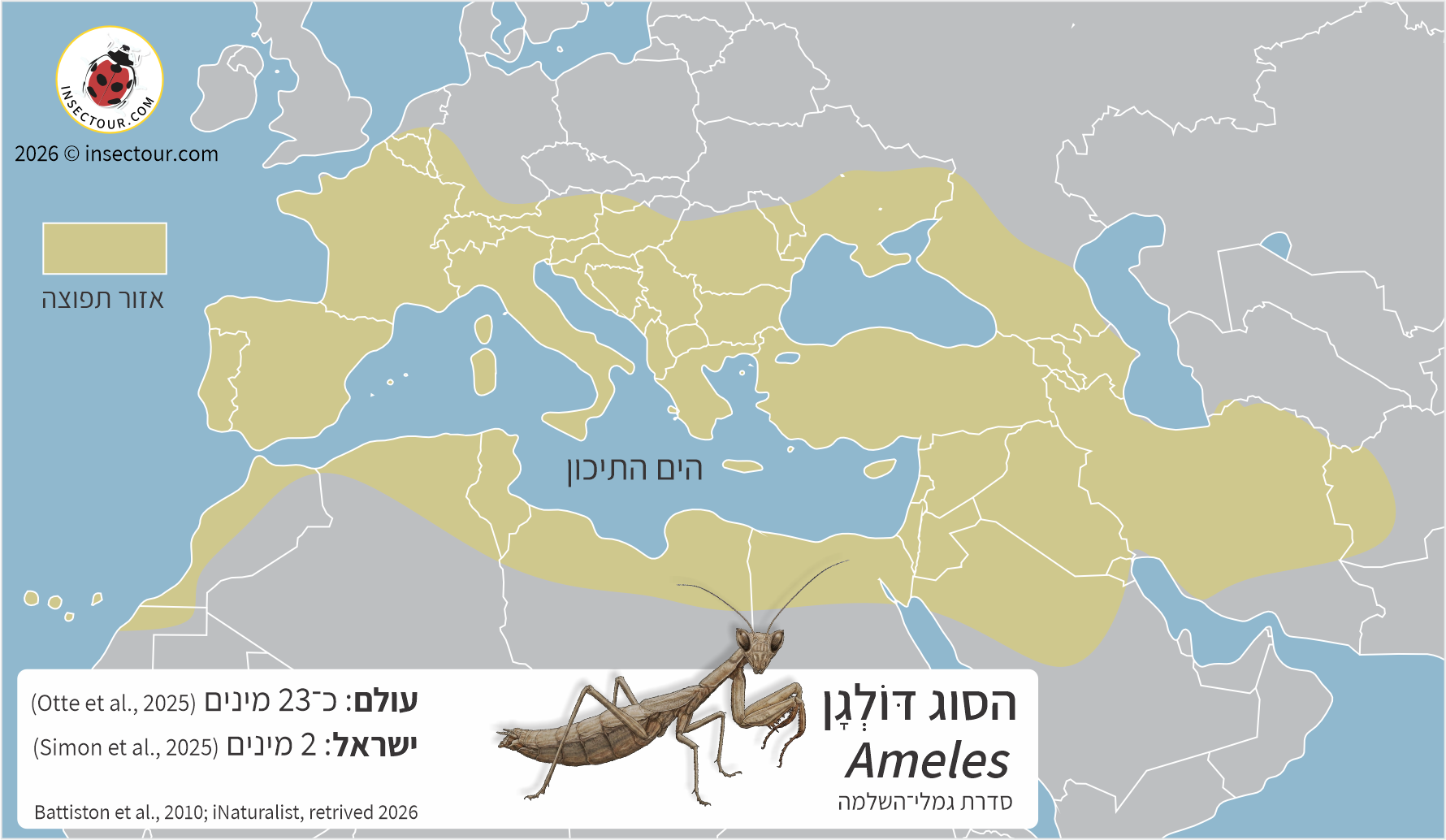
בהקשר זה, הסוג דּוֹלְגָן (Ameles) ומרבית המינים בשבט Amelini הם חריג: מציגים תפוצה פליארקטית הסובבת את אגן הים־התיכון, כולל צפון־אפריקה ומשתרעת עד למערב אסיה. הידע הנוכחי על הביוגאוגרפיה והאקולוגיה של הקבוצה מוביל להשערה שהיא התגבשה והתגוונה במרחב הים־תיכוני, ולא באזור הטרופי או הטרופי־יובשני.
הקשרים האבולוציוניים בתוך משפחת הדֹולְגָנִיִים (Amelidae) טרם פוענחו במלואם. יחסי הקרבה בין המינים במזרח־התיכון ובצפון־אפריקה, עוד הרבה פחות ברורים. באזורים מסוימים זהות המינים עצמם שנויה במחלוקת. אי־בהירות זו מקשה על מתן מענה לשאלות יסוד: היכן הסוג Ameles התחיל להתפתח ולהתגוון? מתי החל לחדור לבתי גידול מדבריים או לרמות הגבוהות והקרות באסיה? ובכלל, כמה מינים יש בסוג?
הסקירה להלן אינה מתיימרת לענות על כל השאלות הללו, אלא להציג, לראשונה בעברית, תמונת מצב נכון לזמן פרסום הרשומה, וידע עדכני על הסוג והמינים בישראל.
משפחת הדֹולְגָנִיִים
סוגיות טקסונומיות אינן תאורטיות בלבד, אלא משפיעות ישירות על האופן שבו מזהים מינים, לומדים אותם ושומרים עליהם.

גם כיום, לאחר למעלה ממאה ושמונים שנים, משפחת הדולגניים (Amelidae) היא עדיין סוגיה מורכבת בהבנה של ההיסטוריה האבולוציונית של גמלי־השלמה.
כטקסון ברמת מִשְׁפָּחָה (family) השם Amelidae הוצע לראשונה על ידי החוקר האיטלקי גִ'ילְיוֹ-טוֹס (Giglio-Tos, 1919). עם זאת, במשך עשורים רבים הקבוצה הוחשבה כתת־משפחה בתוך משפחת הגְּמַל־שְׁלוֹמִיִּים (Mantidae). רק בשנת 2019, בעקבות רביזיה סיסטמתית מקיפה של Schwarz & Roy, בה, בין השאר, פוצלה הגמל־שלומיים למספר משפחות עצמאיות, הדולגניים קיבלו את מעמדם הרשמי כמשפחה נפרדת (Schwarz & Roy, 2019).
מסורתית, המשפחה כוללת שישה סוגים: שלושה בעולם הישן (בשבט Amelini). שלושה בעולם החדש (בשבט Litaneutriini). מחקרים מולקולריים (DNA), מהשנים האחרונות, מראים כי הקבוצה אינה מונופילטית, כלומר היא אינה כוללת אב קדמון משותף אחד ואת כל צאצאיו – הקשר הגנטי בין שני השבטים חלש עד אפסי.
הסוג Ameles (עולם ישן) נמצא כקרוב יותר פילוגנטית לקבוצות אפריקאיות ואירופיות מסוימות. והסוג Litaneutria (עולם חדש) מתמקם בקרבה רבה יותר לקבוצות אנדמיות ביבשת אמריקה. מכך משערים שהמאפיינים המורפולוגיים הדומים בין השבטים נובעים מהסתגלות לסביבות דומות (Ecomorphs) ולא מקירבה גנטית (Svenson & Whiting, 2009). ברשומה זו התייחסות למשפחה היא בהקשר למינים של העולם הישן.
בשנת 2024 התפרסם מחקר פילוֹגנטי שהשווה מספר מינים של דולגניים משלושה סוגים, מרביתם מאירופה, כמה מצפון־אפריקה ואחד מישראל. המחקר אישש במידה רבה את תקפות המיון המורפולוגי המסורתי של המינים של העולם הישן. כמו כן נמצא שהמינים בסוגים Ameles, Pseudoyersinia ו־Apteromantis משתייכים לקבוצה אבולוציונית אחת – חולקים אב קדמון משותף; למרות הבדלים מורפולוגיים ניכרים בינם. המחקר תמך במונופילטיות של השבט Amelini וערער על תקפות הסוג Pseudoyersinia – מאחר שמיניו לא התקבצו לאשכול (clade) אחד, אלא היו מפוזרים על פני העץ הפילוגנטי. לאור זאת, במחקר הומלץ לעדכן את המיון הטקסונומי כדי לשקף במדויק את הקשרים האבולוציוניים (Luchetti et al., 2024).
לפי מחקרם של (2009) Svenson & Whiting, השושלות המובילות לקבוצות הים־תיכוניות המודרניות, וביניהן Amelini החלו להתפצל בסוף האוֹלִיגוֹקֵן, לפני כ־28 מיליוני שנים. במהלך תקופת המִיוֹקֵן (לפני כ־20 עד 15 מיליוני שנים), עברה הקבוצה תהליכי התמיינות והסתגלות משמעותיים. המיוקן התאפיין במגמות עולמיות של התייבשות והתפשטות של שטחים פתוחים. המאמר מציין כי בתקופה זו שושלות אלו עברו התאמה אבולוציונית לחיים קרוב לקרקע. התפוצה הרחבה סביב אגן הים־התיכון מעידה על כך שהקבוצה הייתה קיימת באזור והתפשטה הרבה לפני שאגן הים־התיכון קיבל את צורתו ואקלימו הנוכחיים (Svenson & Whiting, 2009).
בעוד שהתשתית האבולוציונית הונחה במיוקן, האקלים הים־תיכוני המוכר לנו כיום התגבש מאוחר יותר, במהלך הפְּלִיּוֹקֵן וההפְּלַיְסְטוֹקֵן המוקדם והתייצב סופית לפני כ־3.2 עד 2.3 מיליון שנה. יתכן שגם לארוע המסיני (לפני כשישה מיליוני שנים) – במהלכו נסגר מֵצַר גיברלטר לזרימת מים והים־התיכון התייבש כמעט לחלוטין – תרומה חשובה להתפשטות מינים מהיבשות לאיי הים־תיכון ולמעבר יבשתי של מינים בין אירופה לאפריקה (Villani, 2020). ניתן להעריך כי קבוצת ה־Amelini 'מצאה' בשינויים אלו הזדמנות אבולוציונית, שהובילה להסתגלות מוצלחת לבית גידול משתנים ומתהווים.
אחד התרחישים האפשריים הוא שהשבט Amelini התפתח במקביל להתייבשות אזורית של אגן הים־התיכון, כאשר היערות העבותים (באירופה) נסוגו ובמקומם הופיעה צמחייה עשבונית ויובשנית (בתות וערבות), שהחליפה את הכיסוי היער הישן (Suc, 1984). מניחים שהקבוצה עברה 'קרינה אבולוציונית' (radiation) בתוך נישות אקולוגיות שנוצרו ובודדו לסרוגין, תוך כדי התגבשות האקלים הים־תיכוני (Villani, 2020). Agabiti et al. (2010) מציעים שהסוג Ameles החל להתגוון במערב אגן הים־התיכון, אזור המגרב / צפון־אפריקה וחצי־האי האיברי. משם החלה התפשטות מזרחה לאורך חופי הים־התיכון.
מינים כמו דולגן חד־עין מייצגים התפשטות מזרחית. היווצרות חלק מהמינים בסוג דולגן נבעה מבידוד על איים או בתת־אזורים גאוגרפיים; תנאים שהובילו להתמיינות מקומית (אנדמיות) (Agabiti et al., 2010).
הסוג דּוֹלְגָן
לסוג דולגן הסטוריית מחקר פתלתלה, וטקסונומיה שעדיין לא פוענחה במידה מספקת (Battiston et al., 2025). תוצאות המחקר של Luchetti et al. (2024) מדגישות את המורכבות של הפילוגנטיקה של הסוג במזרח־התיכון. בהכללה, אבחנה בין חלק מהמינים בסוג עשויה להיות קשה מאוד; בין השאר גם מתוך שונות מורפולוגית בתוך המינים עצמם; למשל: אורך גוף, אורך יחסי של איברים כמו חזה קדמי, כנפיים וגם וריאציות מבניות באיברי רבייה של הזכרים (בטקסונומיה איברי רבייה מהווים אמצעי זיהוי אמין למדי) (Obertegger & Agabiti, 2012). מקרה כזה מדגיש את החשיבות של מחקר אינטגרטיבי2מחקר אינטגרטיבי משלב טקסונומיה מורפולוגית קלאסית (בחינת מבנה הגוף, העיניים והכנפיים) עם השוואות מולקולריות (DNA). הצלבת הנתונים מאפשרת זיהוי מדויק של מינים, הפרדה בין שושלות אבולוציוניות שנראות דומות חיצונית (אבולוציה מתכנסת) וקביעת קרבה גנטית אמיתית. גישה זו מתקנת טעויות עבר ומבססת סיווג מדעי אמין ומעודכן.המשלב כלים מולקולריים (השוואה מולקולרית – DNA) עם טקסונומיה מורפולוגית קלאסית (Wieland et al., 2014).
נכון להיום, מוכרים בסוג דּוֹלְגָן 25-23 מינים (Otte et al., 2026). זהו הסוג העשיר ביותר במינים במרחב אירופה (Luchetti et al., 2024), הן בתוך משפחת הדּוֹלְגָנִיִּים והן בהשוואה לייצוגן של משפחות גמלי־שלמה אחרות באגן הים־התיכון. מרבית המינים מאכלסים את המדינות והאיים הסובבים את הים־התיכון, וחלקם נחשבים למינים אנדמיים. גבולות התפוצה של הסוג נמתחים מהאיים הקנריים במערב (שם כל המינים בסוג אנדמיים) ועד לחצי־האי ערב ואפגניסטן במזרח.
המאפיינים של הסוג כוללים: אורך גוף קטן עד בינוני – 35-25 מ"מ, הזכרים ארוכים יותר מהנקבות. צמצום כנפיים – זהו מאפיין משפחתי ויתכן שזו תכונה אַפּוֹמוֹרְפִית (מקורה באב קדמון משותף ומיוחדת לקבוצה). לנקבות כנפיים קצרות המגיעות עד פרקי הבטן 3-2; הן לא מעופפות. במרבית המינים לזכרים כנפיים ארוכות מהבטן, אולם תעופתם חלשה ונמוכה. איברי מין זכריים – מבנים ייחודיים באיברי הרבייה מאפשרים לבדל את המשפחה, מציגים שונות מורפולוגית גבוהה בין המינים וגם בתוך המין. ככל הנראה עדות לשושלת עתיקה שעברה תקופות של התגוונות מהירה בתגובה לקיטוע גיאוגרפי באגן הים־התיכון (Villani, 2020).
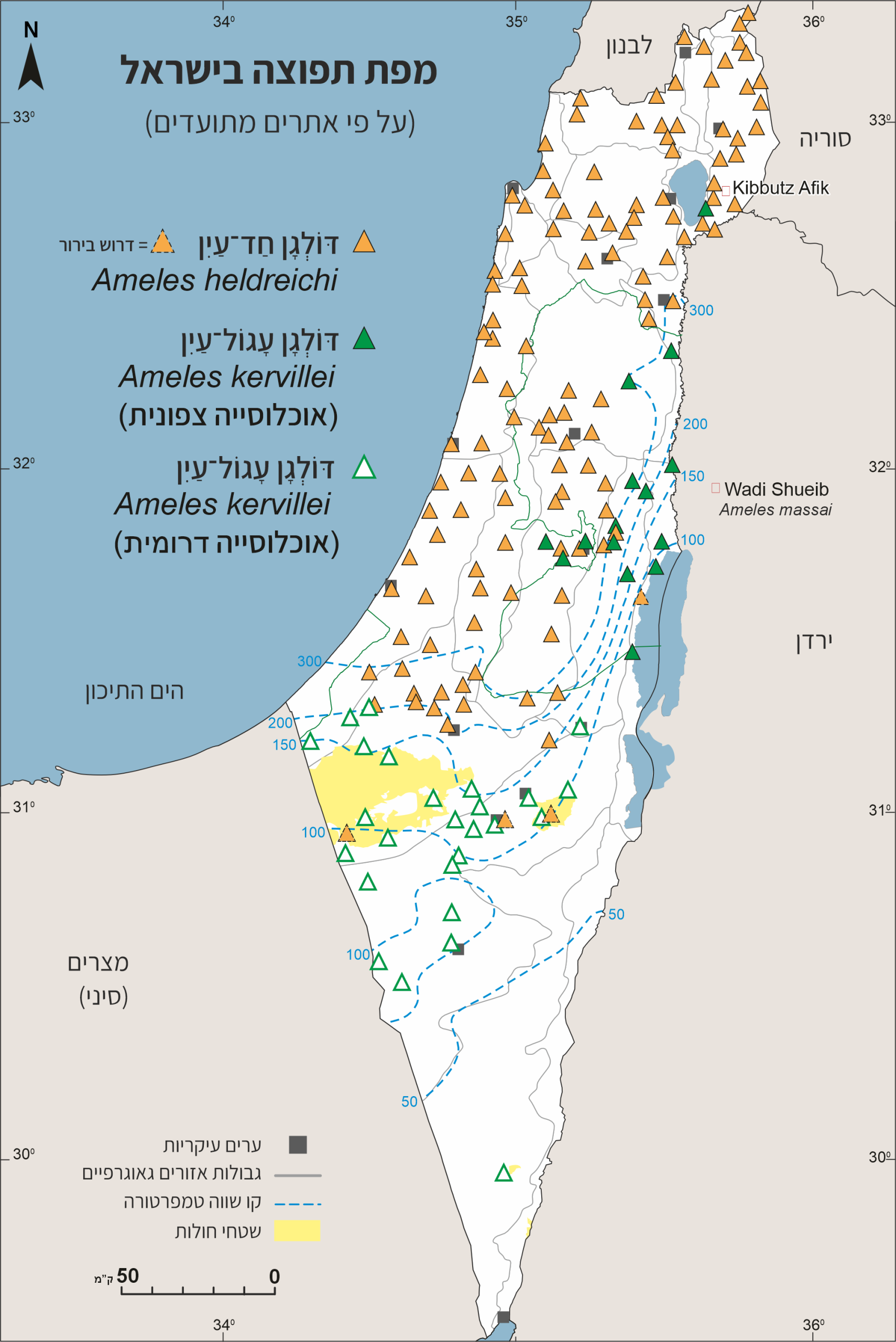
בישראל הסוג מיוצג על ידי שני מינים: דּוֹלְגָן חַד־עַיִן Ameles heldreichi (אָמֵלֶס הֶלְדְרַייכִי) ודּוֹלְגָן עָגוֹל־עַיִן Ameles kervillei (אָמֵלֶס קֶרְוִוילִי). במינים אלו מתקיימות אוכלוסיות שעשויות לייצג פֶנוֹטִיפִּים (פנוטיפ – ביטוי פיזי של תכונה גנטית המושפעת מפעולות גומלין בין גנים לבין הסביבה); אולי בעתיד יסתבר שמדובר במינים נפרדים.
תעופה והפצה

נקבות שאינן מעופפות ותעופה חלשה של הזכרים עשויים להוות עדות להסתגלות לחיים בצמחייה עשבונית דלילה. אף שתעופה משרתת מטרות חיוניות כמו איתור בני זוג, הפצת המין וגמישות בניצול משאבים, היא גם מגבירה משמעותית את החשיפה לטורפים. עבור הנקבות, תעופה גלויה בבית גידול עשבוני פתוח כרוכה בסיכון דור ההמשך; לפיכך, סביר שפועל עליהן לחץ ברירה המרסן יכולות תעופה, מה שהוביל, במהלך האבולוציה שלהן, להתקצרות הכנפיים3לחץ ברירה לטובת כנפיים קצרות מעלה את שכיחותם של פרטים אלו באוכלוסייה, עד שהופכים הם לרוב המכריע בקרב הדורות הבאים. – מצב הכופה מוגבלות מרחבית ניכרת.
עבור הזכרים, התעופה מאפשרת מעבר מהיר בתוך ובין בתי הגידול. התעופה מאפשרת גם איתור נקבות רחוקות, כנראה באמצעות מעקב אחר פרומונים. יכולת זו מעבירה את נטל הפצת המטען הגנטי אל הזכרים, ומסייעת בערבוב גנים בין אוכלוסיות מרוחקות (Roff, 1990). בהקשר זה, הדּוֹלְגָן מהווה דוגמה לאופן שבו לחצי ברירה מנוגדים מעצבים במשולב את אורך הכנפיים, את יכולת התעופה ואת הדינמיקה הגנטית של המין.
לצד ברירה לאובדן תעופה או לתעופה חלשה, הגוררת בידוד גנטי ויצירת אוכלוסיות או מינים אנדמיים, אפשר לראות סתגלנות נוספת, המאפשרת להתגבר על מגבלות הפצה. דוגמא לכך ניתן לראות במין Ameles spallanzania. בשנים האחרונות מין זה מתפשט באירופה צפונה, מעבר למרחב הים־תיכוני החם. הרחבת התפוצה כנראה מושפעת מהאדם (הפצה לא מודעת של תיקי ביצים, כנראה דרך מסחר בצמחי משתלות). סוברים שיש לה קשר הדוק גם לשינוי אקלים, המאפשר התבססות בבתי גידול שבעבר היו קרים עבור התבססות המין (Battiston et al., 2020; Székely et al., 2025).


דּוֹלְגָן חַד־עַיִן Ameles heldreichi
דגם התפוצה כִּתְמִי (לא רציף) ומשתרע בעיקר על דרום־אירופה, דרום הבלקן, טורקיה, יוון, קפריסין, ישראל, ירדן, סוריה, לבנון וצפון־אפריקה.
התפוצה בתחומי ישראל משתרעת ממרומי רכס החרמון ומדרימה, לאורך מרכז הארץ ומישור החוף, עד הנגב הצפוני. עדויות ספורדיות מצביעות על אפשרות שקיימת אוכלוסייה נוספת בנגב המרכזי, שם נדרש סקר תפוצה יסודי שלא נעשה עד כה (Simon et al., 2025).
ההיסטוריה של המחקר

המין תואר תחת השם Ameles heldreichi בשנת 1882 על ידי האנטומולוג החובב השוויצרי פֿוֹן וַאטֶנְוִיל (Brunner von Wattenwyl), ונקרא לכבודו של הבוטנאי הגרמני Theodor von Heldreich שאסף ביוון את הפרט על פיו תואר המין.
הידע על הסוג בלבנט ובישראל מגיע בעיקר מהמין הזה, אולם ההיסטוריה הטקסונומית של המין בלבנט מורכבת. האזכור הראשון של Ameles heldreichi מהאזור שלנו מגיע מהחוקר האיטלקי גִ'ילְיוֹ-טוֹס (E. Giglio-Tos). בפרסום משנת 1893 הוא מתאר אוסף של פרטים שהגיעו אליו מהאוסף של האורניתולוג (חוקר עופות) האיטלקי אנריקו פֶֿסְטָה (E. Festa) שביקר באזור הלבנט בשנת 1893. לפי ג'יליו-טוס באוסף הוא מצא פרט זכר לא בוגר וחסר חלק מהבטן ועוד כמה פרטים מלבנון, גולן וחיפה, שכנראה משתייכים לאתו המין, ללא סטטוס זוויג וגיל. ג'יליו-טוס הסתמך בהגדרות על המפתח של פון ואטנוויל. מאוחר יותר, בשנת 1915 הוא חוזר בו חלקית מההגדרה ומתאר את הפרט מהגולן כמין אחר, חדש למדע – Ameles syriensis.
אזכור שני של Ameles heldreichi, מארץ־ישראל מופיע במאמר של בָקְסְטוֹן (שאסף את הפרטים) ואוּבָרוֹב (שהגדיר אותם במוזיאון הטבע בלונדון). ההגדרה של אוּבָרוֹב כ־Ameles heldreichi נעשתה על בסיס השוואה מורפולוגית לפרטים באוסף במוזיאון הטבע בלונדון, אולם המאמר לא מספק מידע על הפרמטרים ששימשו להשוואה (Buxton & Uvarov, 1923). קביעה זו משמשת עד היום לאישור נוכחות המין בישראל.
נחזור ל־Ameles syriensis שתואר, כאמור, בשנת 1915 על פי פרט יחיד, נקבה שנאספה (1893) באזור פִיק (אפִיק) ברמת הגולן. התיאור שניתן על ידי ג'יליו-טו שטחי. הוא אינו מציג השוואה ברורה, חד־ערכית כנגד דולגנים אחרים שנאספו על ידי פסטה ופרטים מאוספים אחרים באירופה.
במסגרת המחקר על גמלי־השלמה של ישראל נעשתה השוואה מורפולוגית, כולל איברי רבייה של זכרים, בין פרטים מאוכלוסיות שונות בישראל, החל משיא החרמון, רמת הגולן, מרכז ארץ וצפון הנגב. ממצאי ההשוואה לא הצביעו על הבדלים מובהקים בין אוכלוסיית החרמון והגולן לאוכלוסיית אחרת בישראל – למעט שונות טבעית בין אוכלוסיות ובין פרטים באותה אוכלוסייה – שונות צפויה לאור המגוון המורפולוגי באיברי הרבייה. רצף האוכלוסיות מהחרמון ועד לנגב (אין בידוד גאוגרפי או אקולוגי) מחזק את ההנחה שבישראל מין אחד בלבד והשוני בין האוכלוסיות שנבדקו מבטא שונות פנוטיפית (Simon et al., 2025).
בשנת 2026 פורסם מחקר שעסק בדולגן Ameles spallanzania כמין פולש בחצי־האי קְרים. המחקר ערך השוואות מולקולריות גם בין אוכלוסייה מקומית של דולגן חד־עין לפרטים מוזיאונים באירופה שנאספו מהבלקן, מישראל ולבנון. המחקר המולקולרי הראה כי המרחק הגנטי בין הפרטים מקְרים, לאלו מיוון ובולגריה, מזערי (עד 1.71%). מנגד, המחקר הראה כי המרחק הגנטי בין האוכלוסייה בקְרים לבין הפרטים מישראל ולבנון משמעותי יותר (עד 3.97%) – לטענת המאמר אלו הם ערכים גבוליים המשקפים בידוד גאוגרפי ממושך בין אוכלוסיות מזרח הים־התיכון לאלו של הבלקן וקְרים (2026 ,Govorov).
מכאן עולה שאלה מהותית: האם שונות זו מרמזת על היתכנות לתת־מינים או אולי על היות האוכלוסיות בקרים ובלבנט מינים נפרדים? מאחר וישראל מצויה בשוליים המזרחיים של התפוצה העולמית, לא מהנמנע שבלבנט מדובר במין נפרד מהאוכלוסייה באירופה. כדי להבין האם בישראל מתקיים מין שונה מאשר בדרום־אירופה והבלקן נדרש מחקר אינטגרטיבי4מחקר אינטגרטיבי משלב טקסונומיה מורפולוגית קלאסית (בחינת מבנה הגוף, העיניים והכנפיים) עם השוואות מולקולריות (DNA). הצלבת הנתונים מאפשרת זיהוי מדויק של מינים, הפרדה בין שושלות אבולוציוניות שנראות דומות חיצונית (אבולוציה מתכנסת) וקביעת קרבה גנטית אמיתית. גישה זו מתקנת טעויות עבר ומבססת סיווג מדעי אמין ומעודכן. שיכלול אוכלוסיות גם ממזרח אגן הים־התיכון ודרום־אירופה (Simon et al., 2025).


אקולוגיה

בישראל בוגרים נראים בעיקר בין מרץ לנובמבר. המין מקיים בשנה שני דורות עיקריים, אולם לא בכל האזורים. הדור שהגיח בסתיו עובר את החורף בדרגות הנימפה ומתבגר מהר בתחילת האביב. דור שהגיח בקיץ מתרבה בשלהי הקיץ. באתרים בהם הטמפרטורה בחורף קרובה לאפס מעלות מתקיים דור אחד בשנה. באזורים אלו ובמיוחד בחרמון (שם החריפה נעשית בתיק הביצים) הבוגרים הולכים ונעלמים במהלך הקיץ והדור הבא מופיע רק באביב.
ניתוח פעילות חודשי (מרכז הארץ) בעזרת שיטות לכידה שונות בשדה, מראה שפעילות חד־עין באזור הים־תיכוני מושפעת מהטמפרטורה היומית. בעונות הקרות עיקר פעילותו ביום ובעונות החמות חל מעבר לפעילות לילה ובוקר מוקדם (בודנהיימר, 1953).
שכיח בבתי גידול שטופי שמש. שוהה קרוב לקרקע, בין ועל עשבוניים ושיחים נמוכים. עשוי להימצא בישובים כפריים ובשולי מרחבים אורבניים, הנושקים לשטחי בור עשבוניים, שם עשוי להגיע לגינות נוי וחצרות המשופעות בצמחייה נמוכה וצפופה.
תיקי הביצים קטנים (אורכם כ־10 מ"מ), מוטלים לרוב מתחת לאבנים, בסדקי סלע, או בשקעים בשולי סלעים, במפנה מוצל. התיקים פחוסים מעט ומתמזגים היטב עם טקסטורת המצע וצבעו.
סיכון: הנפוץ והמוכר ביותר מבין שני המינים המקומיים. המגמה הכללית של קיטוע בתי גידול ושינוי יעוד של קרקעות מעמידה את המין מול אתגר ענק של בידוד רבייתי והיווצרות אוכלוסיות מבודדות ורגישות העשויות לפתח רגישות לשינויים פתאומיים ולא צפויים.


תובנות

הסוג דולגן משקף מסע בזמן האבולוציוני ובמרחב הגאוגרפי, מסע שבמהלכו שינויים אקלימיים, גאולוגיים וגאוגרפיים דחפו לגיוון ועיצבו אוכלוסיות למינים אנדמיים. במהלך מסע זה הדולגניים הגיעו גם למדבריות נידחים ולבתי גידול הררים – בתי גידול המתאפיינים בתנאי אקלים המוכרים לדולגניים כבר מאותה הסתגלות מוקדמת לאקלים הים־תיכוני היובשני.
אף שמשפחת הדולגניים (Amelidae) זכתה לתשומת לב מחקרית לא מבוטלת, היחסים הפילוגנטיים בין הסוגים והמינים השונים נותרו עמומים. הקושי המרכזי נובע מהיעדר מבט מולקולרי כולל: בעוד שהמורפולוגיה (מבנה הגוף) מתקשה לספק סימני זיהוי חד־משמעיים לאפיון המינים – במיוחד לאור הדמיון הרב בין המינים והשונות התוך־מינית – חסר מידע על איברי הרבייה ומידע גנטי רחב שיאמת את תקפות המינים ברמה הגלובלית והקשרים ביניהם.
ברמה המקומית הסוג דולגן מספק לא פחות עניין ושאלות הדורשות תשובות. כמה מינים יש בישראל ובלבנט? האם האוכלוסיות המקומיות מייצגות תהליך קדום או מתרחש של בידוד גאוגרפי ואולי גנטי?
הפער במחקר המולקולרי מדגיש את הצורך הדחוף בהעשרת האוספים המדעיים בדגימות טריות ושלמות. עד כה, מחקרים על הסוג נאלצו להסתמך, לא פעם, על פרטים מאוספים היסטוריים בני עשרות שנים ויותר; פרטים אלו נמצאים לעיתים קרובות פגומים או שאיכות ה־DNA המופק מהם ירודה, מה שפוגם בהשוואות מהימנות וניתוחים גנטיים מודרניים. רק שילוב בין איסוף שדה עקבי, השוואות מורפולוגיות מתקדמות וטכנולוגיות ריצוף מתקדמות יאפשר לנו לפענח את סיפורה האבולוציוני של המשפחה.