
הסיפור שלנו מתחיל, כמו באגדות, במקרה. השנה: 1798, המקום: פריז, צרפת. בחור צעיר (בקושי בן 21), בוטנאי מוכשר בשם סָבִינְיִי (J. C. Savigny), קיבל הצעה שאי אפשר לעמוד בפניה, להצטרף, על תקן של זואולוג, לצוות משלחת המחקר של מוזיאון הטבע בפריז שתצא למצרים המסתורית כמלווה מדעית לצד צבא נפוליאון.
הלוחות האילמים של משלחת נפוליאון
לאחר כארבע שנים, כאחראי על תחום חסרי־חוליות, במהלכן סייר באזור אלכסנדריה והרחיק עם הצבא הצרפתי לאורך חופי צפון סיני ועד עכו, הוא שב למוזיאון הטבע בפריז עם שפע ממצאים חדשים למדע. במסגרת חלקו בפרסומי ממצאי המשלחת, שהופיעו בשנת 1809, בסדרה המונומנטלית: Description de l'Égypte, הוא היה שותף בהכנת עשרות לוחות מאוירים המציגים את הממצאים הזואולוגים.
בשני לוחות הופיעו גם גְּמַלֵּי־שְׁלֹמֹה. לצד איורים של גמלי־שלמה טיפוסיים, בעלי חזה קדמי מוּאֲרָךְ וכנפיים ארוכות, נראים איורים של גמלי־שלמה 'מוזרים': בעלי גוף קצר מאוד ורחב, כנפיים קצרות ורגלי הליכה ארוכות מאוד (ביחס לאורך הגוף) – שונים מאוד מהמודל הטיפוסי של גמל־שלמה מתפלל (Mantis religiosa) המוכר היטב לאירופאים.
בשל עיוורון מתקדם סָבִינְיִי התקשה להשלים את מפעל חייו ובשנת 1812 הוא נטש את הפרוייקט (Audouin, 1826). הנטישה של סביניי הותירה את הלוחות במצב בלתי אפשרי, האיורים נפלאים, אך הציבו בעייה אחת 'קטנה'; לא מצויין בהם היכן נאספו הפרטים וגם לא שמותיהם.
עמיתו החוקר אוּדוּאָן (Audouin) רצה מאוד לתאר את המינים בלוחות והציע עזרה. סָבִינְיִי שבשל מחלתו הפך למתבודד, סירב בקנאות עד מותו (1851) לחלוק באוסף ובתהילה. בשל היעדרו של סביניי הוטלה על אודואן המשימה המאתגרת, לעשות ככל יכולתו לנסות למספר ולזהות את המינים מהלוחות (חסרי־חוליות, בהם חרקים), וזאת ללא היכולת לראות את הממצאים עצמם, וכך להציל חלק מהידע. עבור גמלי־השלמה הוא הצליח, עם עזרתו של לָטְרֵיי, לזהות כמה פרטים כמשתייכים אל הסוג סוסת־השד (Empusa) ועבור כל השאר הסתפק בתיוג זמני: 'גמלי־שלמה'.
אודואן כותב: "המינים המוצגים תחת המספרים 1, 2, 3, 4, 5 ו-6, למרות שהם שייכים לסוג גמל־שלמה, מהווים בבירור קבוצה חדשה, אלא אם כן הם נחשבים לפרטים לא בוגרים: מר לָטְרֵיי (Latreille) אינו סבור כך" (Audouin, 1826). החלק של הזיהוי וההסברים על הלוחות פורסם בנפרד מהלוחות. למרות הנסיונות של אודואן, הידע על היכן נאספו החרקים, מי במצרים (וסיני) ומי בארץ־ישראל1חלק מהממצאים הזואולוגים שנאספו בארץ־ישראל (שנכללה אז תחת המונח 'סוריה') 'אוחדו' תחת הכותרת הכללית של מצרים בפרסום הסופי, שכן מבחינה זואוגאוגרפית המדענים ראו באזור יחידה אחת – de celles des côtes de l'Égypte et de la Syrie (Savigny, 1809). אבד לעד, וגם ההזדמנות של סָבִינְיִי לתאר מינים חדשים למדע הוחמצה.
מפגש מפתיע בלב הישימון
ברקע הסוגיה על הממצאים של סָבִינְיִי, החוקר לֶפֶבְר (A. Lefebvre), שהיה אנטומולוג מנוסה ובעל מעמד במוזיאון הטבע בפריז, הצטרף (כאנטומולוג) בשנים 1830-1829 למשלחת רפואית צרפתית שנשלחה למצרים לחקור את מגפת הדבר. בשנת 1835 לפבר מפרסם את רשמי המסע (צרפתית) למדבר המערבי (בסמוך לנווה המדבר בחריה) וזיכה אותנו, באופן נדיר לתקופתו, במחשבות לא רק על מה שהוא ראה, אלא גם בתובנות מהתצפיות שלו (Lefebvre, 1835).
לפבר ניצל את הזמן הפנוי בשהותו שם (כחודש בין פברואר למרץ) ואת נתיב המסע לעומק המדבר לתצפיות על חרקים. הוא מציין את הניגודיות בין העושר הביולוגי בנאת המדבר לבין העוני הביולוגי בישימון שמסביבה. במהלך הסיורים הוא פגש 'פנים אל פנים' גמלי־השלמה מוזרים שטרם ראה מעולם. הוא כותב על המפגש הראשון שלו עם גמל־השלמה:
"לאחר יום וחצי של הליכה, מה גדולה הייתה הפתעתי כאשר, בין שברי הקונכיות שמהן אספתי דגימות מפוארות, המופקדות כעת בגלריות המוזיאון, בין המאובנים הללו שרמסו הגמלים שלנו, וביניהם, אך ללא תקווה רבה, עדיין חיפשתי כמה חרקים, מה היה תמהוני, אני אומר, לראות נע באיטיות מעין גמל שלמה קטן בעל גוף מוצק וקומפקטי, חסר כנפיים או כמעט חסר כנפיים, ונראה כאילו הוא מתבונן בחפירות הקטנות ביותר באדמה, כאילו מחפש טרף!"
לפבר השקיע שעות בשטח מאתגר אקלימית בתצפיות על התנהגות, תנועה ויחסי גומלין של החרקים עם סביבתם. הוא התרשם מאוד מעצם קיומם של גמלי־השלמה המוזרים במרחב הלוהט ו"לכאורה חסר החיים" ("semble dépourvu de vie"). מתוך המפגש עם גמלי־השלמה הוא מעלה כמה תובנות מתקדמות (לאותה תקופה) המעידות על ראייה אקולוגית רחבה, מעבר לטקסונומיה יבשה. יש לזכור שהמאמר נכתב כ־24 שנים לפני פרסום 'מוצא המינים' של צ'ארלס דרווין (1859). הידע הדומיננטי בזמנו היה ה'בריאתנות' (הטבע נברא מושלם וקבוע). יש לזכור גם שמושגים מודרניים כמו: אקולוגיה, אבולוציה, אדפטציה ויחסי גומלין לא היו קיימים בזמנו.
א. לפבר התפעם מכך שגמלי־שלמה אלו מותאמים בגוון גופם לקרקע (חום על אדמה, כסוף על גיר). הוא תהה האם הם משנים את צבעם בהתאם למשך שהותם בשטח, אך ציין כי לדעתו "יכולת זיקית" זו (שינוי צבע דינמי) טרם נצפתה בקרב חסרי־חוליות. את ההתאמה המוחלטת הזו הוא תפס כ'כוונת הטבע' שנועדה להעניק להם הגנה מפני הטורפים המעטים בישימון.
ב. הוא הבחין כי גמלי־השלמה הללו הם החרקים היחידים בישימון הראויים לשמש כטרף לאויביהם. מכך הוא הסיק שהטבע 'התאים' את כסותם לצבע הקרקע כדי להקל עליהם לחמוק מהטורפים, עד כדי כך שכמעט בלתי אפשרי להבחין בהם, במיוחד כשהם במצב של חוסר תנועה.
ג. לפבר הוטרד עמוקות מהשאלה כיצד חרק שכל איבריו (רגלי הטרף) מעידים עליו שהוא חי על "ביזה ומלחמה" [טורף מובהק], מסוגל להתקיים באזור שומם לחלוטין שאין בו צמחייה, שרידי צמחייה או חרקים אחרים שישמשו לו למאכל. הוא הגדיר את תנאי המחיה האלו כ'בלתי ניתנים להסבר' במסגרת חוקי הטבע המוכרים לו.
ד. הוא שלל את ההנחה שגמלי־שלמה אלו נודדים או נישאים בסופות אל האזורים המעובדים כדי להזין את עצמם ושבים למדבר. על סמך התנהגותם הבודדת והשקטה, הוא משתכנע כי המדבר הקיצוני הוא משכנם הקבוע והבלתי משתנה, וכי הם אינם עוזבים אותו לעולם (Lefebvre, 1835).
באחת מהערות השוליים למאמר, לפבר מחדד את התפיסה האקולוגית שלו ומציג לכאורה יכולת מרשימה של הסקת מסקנות רחבה. יתכן שעל סמך היכרותו עם תיאורי האזור וידיעתו כי בפנים היבשת של סוריה ולבנון קיימים שטחים נרחבים בעלי אופי יובשני, הוא מוצא את המכנה המשותף בין המדבר המצרי הקיצוני לבין אזורי הלבנט. הוא מבין שהזיקה האקולוגית של הסוג היא לתנאי אקלים יובשני ושטחים פתוחים ולא לשממה הקיצונית כשלעצמה, ומנסח תובנה הגיונית וכללית לגבי העדפת בית הגידול שלהם: "עם זאת, עלינו להאמין שחרקים אלה אינם מאכלסים אך ורק את המקומות שבהם צמחייה בלתי אפשרית… אני חושב שאנו יכולים, ללא חשש לטעות, להאמין שגמלי־השלמה הללו מאכלסים מקומות יבשים יותר מאשר מקומות מעובדים".

אתגר טקסונומי

אף שהסוג מדברן מוכר למדע כבר למעלה ממאתיים שנים, הטקסונומיה שלו נותרה סבוכה. מרבית המינים בקבוצה זו תוארו במהלך המאה ה־19 וראשית המאה ה־20, בתקופה שבה התיאורים המדעיים היו לרוב שטחיים והתבססו על מאפיינים מורפולוגיים חיצוניים בלבד. כיום מקובל שהבדלים חיצוניים בלבד אינם מהווים תמיד סממן מובהק להפרדה בין מינים. קושי נוסף נובע מכך שמינים רבים הוגדרו על בסיס פרטים בודדים, לעיתים פגומים או כאלו שנאספו בשלב הנימפה. כך יתכן שזכרים ונקבות מאותו המין תוארו כמינים נפרדים.
גם כיום, חסר ידע בסיסי על המורפולוגיה של איברי הרבייה בסוג – כלי מרכזי בזיהוי טקסונומי מודרני – וטרם בוצע ניתוח קלדיסטי מולקולרי (DNA) מעמיק שישפוך אור על יחסי הקרבה האבולוציוניים בתוך הסוג (Battiston et al., 2010).
מורפולוגיה
גוף המדברן קצר ופחוס. אורכו של בוגר (מקדמת הראש עד קצה הבטן) לרוב לא עולה על 36 מ"מ במינים גדולים. במינים קטנים, לרוב לא עולה על 15 מ"מ. רגלי ההליכה ארוכות ומותאמות לריצה מהירה על קרקע חשופה. צידוד הראש, המוכר בגמלי־שלמה חובבי צמחים, מוגבל – אפשר שזו הסתגלות לציד על הקרקע, של טרף המתגלה בטווחי מרחק קצרים מאוד.
איבר שמיעה, השכיח בגמלי־שלמה מעופפים, חסר. הכנפיים אינן מותאמות לתעופה, אולם מערכת שרירי הכנפיים נותרה פעילה. מצב התומך בהשערה שהקבוצה התפתחה מאב קדום מעופף (Schwarz & Roy, 2019), מכאן שזו עדות לאובדן תעופה מִשְׁנִי. שרירי הכנפיים מאפשרים רק פרישת כנפיים המשמשות כעת למצג הרתעה ובחיזור (על כך בהמשך).
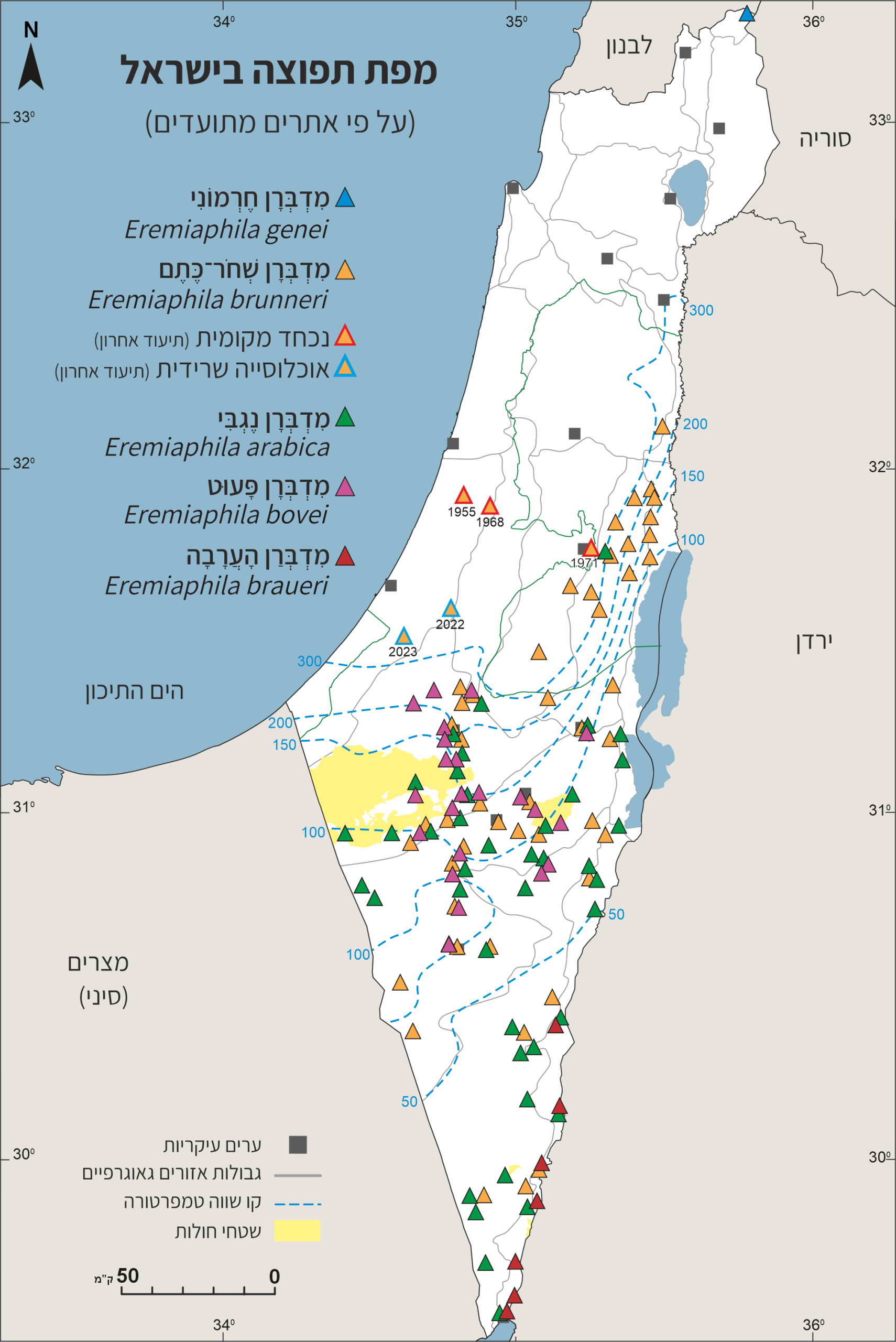
תפוצה ועושר המינים

פַאוּנָת גְּמַלֵּי־הַשְׁלֹמֹה של ישראל מונה כ־33 מינים (Simon et al., 2025). במהלך מיליוני שנים התפתחה במזרח־התיכון פאונה מקומית ממינים שמוצאם מאזורים הקשורים קשר הדוק לאזורנו או שהיה להם קשר כזה בעבר הקדום. מרביתם שוכנים רק או בעיקר באזורים צחיחים, לרוב מדבריים. חלקם אנדמיים לישראל או לישראל והשכנות הקרובות. על האזורים הללו נמנים: צפון־אפריקה (הסהרה), מזרח־אפריקה (האזור הסודני / אתיופי), חצי־האי ערב, דרום־מזרח אסיה ואגן הים־התיכון – אזורים המתאפיינים באקלים טרופי, טרופי-יובשני, מדברי, ממוזג-ים־תיכוני.
הסוג מדברן שוכן באזורים צחיחים ויובשניים למחצה, המתפרסים בדרום האזור הפליארקטי, מחופי מערב אפריקה ועד הודו (ראו מפה). מתוך כ־64 מינים (Otte et al., 2025), 47 תועדו מאזור אגן הים־התיכון בכללותו. כ־38 רק בצפון־אפריקה וכ־39 במזרח־התיכון. כחמישה מינים מוכרים ממרכז אסיה (ארמניה, אירן, אזרביג'ן, אפגניסטן, פקיסטן, הודו). מספר המינים הגדול ביותר: כ־29, מוזכר ממצרים, כ־15 ידועים רק משם.
עושר כזה של מינים דומים (חזותית והתנהגותית), ובמיוחד שיעור האנדמיות הגבוה, בשטח גאוגרפי נרחב אך מקוטע אקולוגית וטופוגרפית, הוא ככל הנראה עדות להתפתחות והתגוונות אוכלוסיות תחת השפעה של בידוד סביבתי, הנוצר עקב מגבלות ניידות (אי יכולת לעוף) ושפע בתי גידול דומים (המשמרים את המאפיינים של הסוג) אך מנותקים (בידוד גאוגרפי וטופוגרפי) – דגם קלאסי של התמיינות אַלוֹפַּטרית באזורים מקוטעים (Mayr, 1963; Brito et al., 2013; Hewitt, 2000).
במהלך המחקר על גמלי־השלמה של ישראל הובררה הימצאותם של לפחות חמישה מינים בסוג מדברן; אחד בחרמון וארבעה בנגב ובמדבר יהודה. לא מהנמנע שבישראל ימצאו בעתיד מינים נוספים, שישראל מצויה בגבול תפוצתם העולמי, או ה'מסתתרים' בתוך מינים מוכרים (מינים כמוסים) או השוכנים באזורים שלא נסקרו (Simon et al., 2025).




בניגוד לגמלי־שלמה טיפוסיים הצדים ממארב על צמחים, ביום וגם, או בלילה. המדברן הוא פעיל יום משוטט, התָּר אחר טרף מזדמן. פעילות לאור יום בשטח חשוף לגמרי עשויה להיות גלויה לטורפים. מאידך, במדבר קיצוני הזדמנויות הציד נדירות. כאשר עושר וצפיפות הטרף קטנים יחסית, חיפוש אקטיבי אחר טרף, ביום חשוף לעין כל, הוא אסטרטגיה המאזנת בין הסיכון לבין הרווח הפוטנציאלי: השגת מזון בחיפוש פעיל לעומת המתנה פסיבית לטרף אקראי.
בתנאי מדבר אין יתרון לבררנות תזונתית – המדברן יצוד כמעט כל פרוק־רגליים שיזדמן ולא עולה על גודלו שלו. בעיקר חרקים, כולל נמלים שאינן נחשבות טעימות ונמצאו בתכולת הקיבה של מדברן (Roonwal, 1938). Adair (1914) מציין שבשבייה הייתה העדפה לאכילת נמלים (לא מצוין סוג), אבל גם זבובים נאכלו ברצון. הידע על התזונה בטבע נסמך על תצפיות מעטות ויש עוד הרבה ללמוד על מאפייני התזונה של מדברנים בבתי גידול שונים.
בניגוד לשוכני מדבר רבים הנמנעים מתנאי קיצון של חום (פעילים בלילה או בעונה הפחות חמה) המדברן לא 'ברח' הסתגלותית מהקושי, אלא התפתחו בו, במהלך האבולוציה, התאמות התנהגותיות ופיזיולוגיות המאפשרות פעילות בתנאי קיצון ובמיוחד בחום, ייתכן תוך פשרה עם יכולות אחרות, כגון תעופה או הפצה למרחקים. הסתגלות לחום קיצוני מאפשרת הסטת זמני פעילות וחיפוש מזון (לדוגמה: נמלים הפעילות בחום) לשעות בהן פעילות של מתחרים וטורפים מצומצמת.
המדברנים מתחילים פעילות בבוקר המאוחר ומשהים פעילות בימים קרירים. (1923) Buxton דיווח על נימפה של מדברן שנמצאה פעילה בחודש יוני בצהרי יום ליד יריחו, כאשר טמפרטורת האוויר הייתה 32.30C מעלות בצל ונמדדה טמפרטורת הקרקע של 50.80C. החוקר (1969) Délye השווה עמידות להתייבשות וחום בחרקים מדבריים, בהם גם המדברן Eremiaphila monodi מאלג'יריה. הוא מצא שהמדברן מסוגל לשרוד למעלה משעה בחשיפה לטמפרטורה של כ־500C מעלות ואובדן מים משמעותי מתחיל מעל 600C. עמידות המעידה על חדירות נמוכה של הקוטיקולה לנוזלים ועל פתחי נשימה (דרכם אובדן הנוזלים היחסי גדול) המותאמים לתנאי יובש. ייתכן, גם, שרגליים ארוכות מסייעות להרחיק את הגוף מהקרקע הלוהטת.
Zera & Denno (1997) מציינים כי לחרקים מעופפים יש קצב נשימה (respiration rate) גבוה יותר בשל הצורך לחמצן שרירי תעופה מסיביים. בפיזיולוגיה של חרקים, קצב נשימה מוגבר מוביל לרוב לאובדן מים רב יותר דרך הטרכאות (מערכת הנשימה), כך שבעקיפין, חרק שאינו עף ובעל קצב מטבולי נמוך יותר עשוי 'לחסוך' במים.
מינים הרריים הסתגלו לפעילות בטמפרטורות נמוכות. חוקר הזוחלים הֶרְמָן צינר (האוניברסיטה העברית) סייר בלבנון בשנים 1956-1955 ומדווח – במאמר שפורסם במגזין Israel Journal of Zoology – על מדברנים שנראו ונאספו בצפון הלבנון, ברכס קורנת א-סוודא, בגובה 2900 מטרים מעל פני הים (Zinner, 1967). הספרות מציינת שבאזור זה טמפרטורת המקסימום בקיץ היא 100C-150C מעלות ובלילה קרוב לאפס. בשיא החרמון לדוגמה, הטמפרטורה היומית בקיץ לא עוברת את ה־250C מעלות ולרוב היא 190C-220C מעלות.
באזורי תפוצה חמים מדברנים פעילים מרבית השנה. בישראל בוגרים נראים לרוב מאביב מאוחר עד לסתיו מוקדם ויש מינים בהם בוגרים וצעירים נראים מרבית השנה. באזורי תפוצה קרים הפעילות מתרכזת בשלהי האביב ובקיץ והחריפה (ככל הנראה בדִיאַפָּאוּזָה) נעשית בדרגת ביצה, המוגנת בתיק ביצים (ootheca).



גדילה והתפתחות

בשבייה נקבה, הניזונה היטב, עשויה לייצר עד 10 תיקי ביצים במרווחים של לפחות 10 ימים, לרוב יותר. בטבע מספר התיקים קטן בהרבה ומרווחי ההטלה גדולים יותר, כתלות במין ובאיכות התזונה. מהתיק מגיחות 15-10 נימפות. בשונה מרוב גמלי־השלמה, הנעזרים בכוח הכבידה כדי להגיח מהתיק הממוקם מעל הקרקע (על צמח או סלע), נימפות המדברן מגיחות מהקרקע כלפי מעלה.
תזמון גיחת הנימפות (אורכם 4-2 מ"מ) משתנה ממין למין. מתצפיות בשבייה (ללא ציון מינים) מסתמן שיש מינים בהם הנימפות מגיחים יחד ויש כבודדים במהלך מספר שבועות עד מספר חודשים. נראה שהלחות היחסית מהווה גורם המשפיע על תזמון ההגחה, כאשר הזחלים מעדיפים להגיח עם שחר, כשהלחות בשיאה.
על פי דיווחים משבייה (ללא ציון מינים) ההתפתחות מהירה ופרטים הניזונים היטב עשויים להגיע לבגרות לאחר כחודשיים. בטבע תהליך זה ככל הנראה ארוך יותר.
בגמלי־שלמה זכרים מתפתחים מהר מהנקבות, עם דרגת התפתחות אחת פחות מהן. הזכרים מעט קטנים מהנקבות, הכנפיים ארוכות במעט (יחסית לאורך הגוף) והמחושים ארוכים יותר. בשבייה זכרים חיים מספר שבועות והנקבות עד מספר חודשים.
![]() טיפ זיהוי! נימפות של מדברן עשויות להיחשב בטעות לבוגרים, ובוגרים של מינים קטנים לנימפות. כלל אצבע לזיהוי: אצל בוגר של גמל־שלמה מכונף, הכנפיים (בין אם קצרות או ארוכות) חופות מעל הבטן; אצל נימפה, ניצני הכנפיים ערוכים לצידי הבטן ואינם חופפים. בישראל רק המין עַצְבָּנִי עָלוּם, הוא חסר כנפיים באופן ראשוני, בכל הדרגות.
טיפ זיהוי! נימפות של מדברן עשויות להיחשב בטעות לבוגרים, ובוגרים של מינים קטנים לנימפות. כלל אצבע לזיהוי: אצל בוגר של גמל־שלמה מכונף, הכנפיים (בין אם קצרות או ארוכות) חופות מעל הבטן; אצל נימפה, ניצני הכנפיים ערוכים לצידי הבטן ואינם חופפים. בישראל רק המין עַצְבָּנִי עָלוּם, הוא חסר כנפיים באופן ראשוני, בכל הדרגות.












 מדע אזרחי – חשוב לדעת:
מדע אזרחי – חשוב לדעת:
אחד האתגרים במחקר אקולוגי על הסוג מדברן והמינים מקומיים, שהם פעילים בחום ויובש שאינם נוחים לאדם. הנגישות לאוכלוסיות, שחלקן שוכנות באזורים רחוקים, קשה. מנגד חובבי טבע רבים מסיירים ומבקרים כל השנה באזורי התפוצה של המדברנים.
חובבי טבע הנפגשים עם מדברנים (וגמלי־שלמה בכלל) יכולים לתרום להרחבת ההיכרות עם הסוג המרתק הזה. תיעוד מצולם (עם תאריך לועזי מלא ומיקום מדוייק, החיוני להבנת בית הגידול. בצרוף תיאור כתוב על התנהגויות שונות: טריפה, חיזור, הזדווגות, הגנה, הסוואה, בית הגידול, שעות פעילות ועוד. פרסומם במדיות חברתיות (פייסבוק, iNaturalist), יכולים להוות תרומה משמעותית לידע המדעי והציבורי על התפוצה, הביולוגיה והאקולוגיה של המינים בסוג. עבור החוקר שלא יכול להיות בכל מקום בכל זמן, מאות ואלפי תצפיות קטנות הן מכפיל כוח, חלקי פאזל שאפשר לחבר לתמונה רחבה יותר, ממנה ניתן ללמוד הרבה יותר. עבור חובבי הטבע גם תצפית קטנה מעניקה חוויה, לעיתים ייחודית, על קבוצת חרקים מיוחדת – שאינה דורשת הרבה מלבד עיניים טובות וסבלנות, כדי ללמוד עליה דברים חדשים.